Membres du projet


Projet
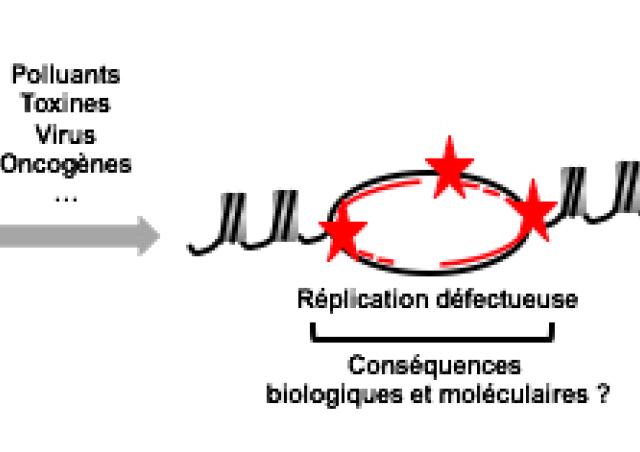
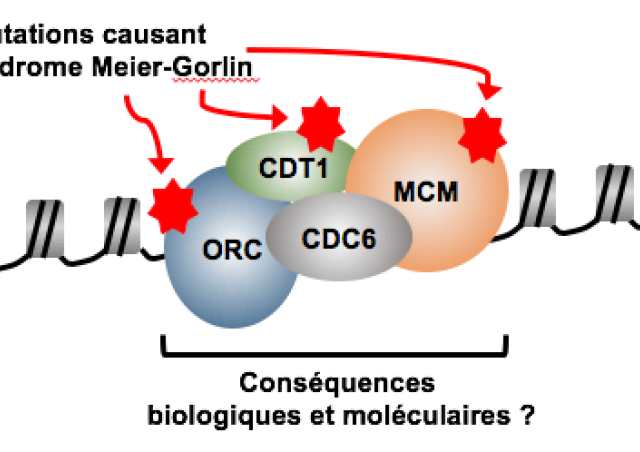
L'initiation de la réplication débute au niveau des milliers d'origines de réplication, distribuées le long de la fibre d'ADN. Les caractéristiques de ces origines restent encore à élucider, aussi bien au niveau des séquences ADN que des protéines retrouvées à ces origines. La première étape est la liaison à la chromatine du complexe de reconnaissance des origines, aussi appelé ORC, qui se compose des sous-unités ORC1-6. La liaison de ce complexe permet d’établir la position des origines de réplication en sortie de mitose, et permet ensuite le recrutement des facteurs essentiels à l'initiation de la réplication comme les protéines CDC6 et CDT1 et le complexe MCM2-7.
Il est admis que le recrutement du complexe ORC à la chromatine, ainsi que l'assemblage des protéines essentielles à l’initiation de la réplication, sont régulés par l’organisation et la nature des marques présentes sur la chromatine. L'équipe a pu montré, par exemple, que l'acétylation des queues d'histone favorise le recrutement du complexe MCM2-7 aux origines de réplication (Miotto et Struhl, 2008 et 2010). Cependant, les liens moléculaires et fonctionnels entre l'organisation de la chromatine et l'activité des origines de réplication restent à être mieux définis.
Pour répondre à cet enjeu, notre équipe a produit une cartographie des régions du génome liées par l'une des sous-unités du complexe ORC, la proteine ORC2 (Miotto et al., 2016). Cette information nous permet actuellement de développer de nouvelles analyses bio-informatiques, moléculaires et cellulaires pour comprendre :
- Quelles sont les caractéristiques de séquence et de chromatine des origines de réplication ?
- Quel rôle joue l'organisation de la chromatine sur l’assemblage et la fonction des protéines d’initiation de la réplication ?
- Quels sont les liens fonctionnels entre l’activité des origines de réplication et les autres fonctions nucléaires ?
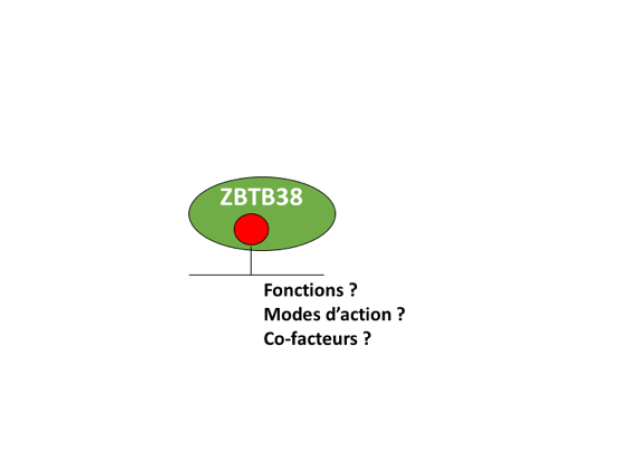
Au cours des dernières années nous avons contribué à démontrer les fonctions inattendues aux origines de réplication des facteurs de transcription NFY (Benatti et al., 2016) et ZBTB38 (Miotto et al., 2014) ainsi que de l'acétylase HBO1/KAT7 (Miotto et al., 2008). Un aperçu vulgarisé de ces travaux est disponible ici.
Nous avons aussi participé à la caractérisation de nouveaux régulateurs du processus de réplication, comme la protéine GNL3 (Lebdy et al., 2023), la protéine de réparation FANCD2 (Fernandes et al., 2021) ou le facteur d'épissage SF3B1 (Rombaut et al., 2024).